Нейрофизиологический и формально-логический базис нейроподобных вычислений
4.1. Системотехнический анализ морфофункциональных схем нейронов, нервных сетей и механизмов "спайковой" активности
Нервная система человека и животных состоит из однотипных клеток, именуемых нейронами, которые связаны между собой разветвленной системой аксонно-коллатеральных связей (рис. 4.1). Главная отличительная черта нейронов - это наличие в невозбужденном состоянии разности электрических потенциалов (потенциал покоя - рис. 4.2) на полупроницаемой мембране и аксонно-коллатеральных связях. Это обусловлено разной концентрацией ионов калия и натрия на внешней и внутренней поверхности мембраны, которая в значительно большей мере препятствует ионам натрия диффундировать внутрь клетки (рис. 4.3).

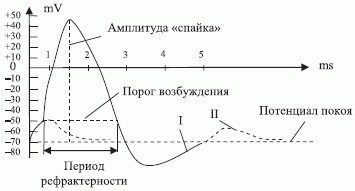
Эта разность потенциалов поддерживается силами поверхностного натяжения мембраны и поэтому определяется ее биофизическими и биохимическими свойствами, которые изменяются под воздействием различных физико-химических раздражителей. В результате изменяется проницаемость мембраны, что приводит к двунаправленной диффузии анионов калия и натрия, которая приводит к изменению потенциала покоя и к генерации одиночного "спайка" с достаточно стабильными амплитудно-временными параметрами и формами (рис. 4.2). Существенно, что диффузия анионов ("калий-натриевый насос") срабатывает ортогонально по отношению к направлению распространения "спайка", амплитуда которого у каждого нейрона стабильна, но у разных нейронов колеблется в пределах 70-90 мВ. Передний фронт "спайка" имеет два характерных участка, разделенных порогом возбуждения, превышение которого и приводит к генерации импульса. Задний фронт "спайка" может быть апериодическим (крива я I) либо затухающим колебательным (кривая II). Непревышение порога возбуждения приводит к локальной деполяризации мембраны нейрона, а после превышения запускается механизм типа "все или ничего". С системотехнических позиций такие стабильные амплитудно-временные параметры отдельных "спайков" говорят о том, что информация о реализуемых нейронами функциях может быть заложена только в параметры "спайковых" последовательностей, что в технике соответствует частотно-импул ьсной или времяимпульсной модуляции, последняя из которых носит аналоговый характер. При этом следует помнить:
- В "спайковых" последовательностях максимальная частота или минимальное время между импульсами регламентируется периодом рефрактерности (см. рис. 4.2), когда нейрон просто не чувствителен к входным воздействиям.
- В нейрофизиологии гораздо важнее идентифицировать участие (неучастие) того или иного нейронного образования в формировании или регулировании той или иной жизненно важной функции, чем количественные соотношения между стимулом и реакцией. Поэтому в нейрофизиологическом эксперименте оценивается не количество переданной по аксону информации, а факт изменения фоновой активности подконтрольного нейрона на тот или иной тип и величину внешнего воздействия. Для этих целей сначала регистрируется фоновая "спайковая" последовательность подконтрольного нейрона и определяются ее статистические характеристики (математическое ожидание и/или дисперсия). После этого на подопытное животное подается внешнее воздействие, регистрируются и оцениваются изменения "спайковой" активности того же нейрона, по которым и делается вывод об участии в формировании реакции организма на поданное внешнее воздействие той области мозга, к которой принадлежит подконтрольный нейрон. При этом оцениваемые количественные соотношения в больш ей степени отражают качественные изменения типа "чем сильнее воздействие, тем сильнее (слабее) реакция" и не более. Механизм действия "калий-натриевого" насоса сводится к следующему. Полупроницаемая мембрана в состоянии покоя препятствует проникновению ионов натрия внутрь клетки, вследствие чего на внешней стороне мембраны образуется "больший плюс", чем на внутренней стороне. В активном состоянии проницаемость мембраны для ионов натрия возрастает в 400-500 раз, а для ионов калия всего в 10-15 раз. Благодаря такой разности в изменении проницаемости мембраны встречная диффузия ионов калия и натрия приводит к локальной деполяризации участка мембраны, когда состоянию "больший плюс" отвечает уже концентрация анионов на внутренней стороне мембраны [55].
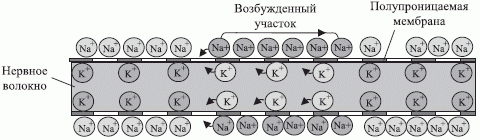
В отличие от коллатералей аксон покрыт миелиновой оболочкой, секционированной перехватами Ранвье (см. рис. 4.1), в результате чего "спайк" распространяется по нему с более высокой скоростью, которая колеблется в пределах 1-100 м/сек в зависимости от толщины миелинового слоя.
Сома нейрона отделена от аксона и связанных с ним коллатера-лей аксонным "холмиком". При этом в электрофизиологической модели Ходжкина - Хаксли [56] аксонному "холмику" отведено центральное место в формировании "спайка", механизм распространения которого по аксону и показан на рис. 4.3. Согласно этой модели аксонный "холмик" имеет самый низкий порог возбуждения на мембране нейрона, а условие трансформации входных нейрохимических воздействий в "спайк" имеет вид:
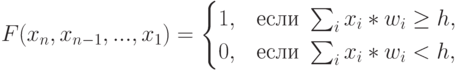 |
( 4.1) |
где 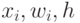 - соответственно входные воздействия на нейрон, весовые коэффициенты "восприятия" входных спайков на мембране нейрона и порог возбуждения нейрона.
- соответственно входные воздействия на нейрон, весовые коэффициенты "восприятия" входных спайков на мембране нейрона и порог возбуждения нейрона.
"Взвешенному" суммированию в (4.1) отвечает нейрохимический механизм передачи возбуждения через синаптическую щель (рис. 4.4 [57]), которая отделяет аксон или коллатерали от мембраны принимающего (постсинаптического) нейрона.
![Структура синапса [3]](/EDI/14_06_16_2/1465856498-31733/tutorial/593/objects/4/files/04_04.gif)
Пресинаптическое волокно (аксон или коллатераль) заканчивается синаптической бляшкой, внутри которой расположены митохондрии и пузырьки размером 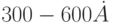 , которые содержат специальные вещества (медиаторы) и скапливаются в одной или нескольких областях синап-тической щели. Существует как минимум два типа синапсов, которые отличаются шириной щели и толщиной постсинаптической мембраны (рис. 4.4-а и 4.4-б соответственно), причем синапсы второго типа принято относить к возбуждающим, так как они характерны для достаточно длинных аксонов и образуемая ими щель в 1,5 раза уже (соответственно
, которые содержат специальные вещества (медиаторы) и скапливаются в одной или нескольких областях синап-тической щели. Существует как минимум два типа синапсов, которые отличаются шириной щели и толщиной постсинаптической мембраны (рис. 4.4-а и 4.4-б соответственно), причем синапсы второго типа принято относить к возбуждающим, так как они характерны для достаточно длинных аксонов и образуемая ими щель в 1,5 раза уже (соответственно  и
и  ). Наличие митохондрий в синаптических бляшках указывает на возможность воспроизводства медиаторов непосредственно в синапсах.
). Наличие митохондрий в синаптических бляшках указывает на возможность воспроизводства медиаторов непосредственно в синапсах.
Синаптические механизмы возбуждения ткани были впервые обнаружены в нервно-мышечных окончаниях, регулирующих (координирующих в пространстве и во времени) процессы сжатия и растяжения мышцы в целом. Исследования возбуждающих синапсов показали [57], что пре-синаптические импульсы неизменно вызывают локальную деполяризацию постсинаптической мембраны. В чисто нейронных синапсах такая реакция именуется возбуждающим постсинаптическим потенциалом (ВПСП), а ее эквивалент в нервно-мышечном окончании - потенциалом концевой пластины (ПКП).
Экспериментально был показан эффект пространственной сум-мации [57], когда несколько синхронных пресинаптических импульсов повышали амплитуду ВПСП, но не длительность локальной деполяризации постсинаптической мембраны.
Одним из главных показателей синапсов является время задержки от момента прихода пресинаптического импульса к бляшке до начала постсинаптической деполяризации, регистрируемой внеклеточно. В зависимости от функционального назначения нейрона и типа подопытного животного время задержки на синапсах колеблется от 0,3 мсек и до 2 мсек, где первое значение характерно для центральной нервной системы млекопитающих и для нервно-мышечных соединений поперечно-полосатых мышц [57].
Длительное время в физиологии синапсов противостояли две теории передачи возбуждения через синапс: более ранняя - электрическая и более поздняя - нейрохимическая. Сторонники нейрохимических механизмов передачи возбуждения через синаптическую щель исходили из наличия медиаторов, которые должны удовлетворять следующим требованиям:
- данное вещество должно присутствовать в достаточных количествах в пресинаптических окончаниях, которые должны включать в себя синтезирующую ферментную систему;
- раздражение пресинаптических нервов должно приводить к высвобождению достаточного количества этого вещества из пресинапти-ческих окончаний и его диффузии через синаптическую щель;
- действие этого вещества на постсинаптическую клетку должно быть идентично синаптическому действию, если само вещество вводится методом микроэлектрофоретической инъекции;
- в области синаптической щели должна существовать инактивирую-щая это вещество ферментативная система;
- при изучении действия фармакологических препаратов методом микроэлектрофоретической инъекции их влияние на синаптическую передачу и на постсинаптическое действие исследуемого вещества должно быть одинаковым.
Всем этим требованиям на момент наиболее успешных исследований синаптических механизмов удовлетворяли только ацетилхолин и нора-дреналин, метаболизм которых и был изучен достаточно полно. Показано [57], что с ростом частоты пресинаптических импульсов выделение аце-тилхолина возрастает в 70 раз по отношению к состоянию покоя, а интенсивность его синтеза увеличивается в 7 раз. При этом были раскрыты механизмы депонирования избыточного ацетилхолина, что в совокупности создает условия для адекватного реагирования синапса на возрастание частоты пресинаптического возбуждения.
Многообразие синапсов с электрической формой передачи возбуждений гораздо шире, чем с нейрохимической. Но во всех случаях передача в таких синапсах осуществляется только в одном направлении и посредством деполяризующего действия электрического тока, который создается либо импульсами, либо синаптическими потенциалами в пре-синаптической мембране [57].
Приведенные данные говорят об аналоговом или как минимум циф-роаналоговом механизме локальной деполяризации постсинаптической мембраны, первый из которых характерен для синапсов с электрической передачей возбуждения, а второй - с нейрохимической. Вместе с тем, они не раскрывают:
- механизмы трансформации локальных деполяризаций сомы и/или дендритов нейронов в "спайковую" активность всего нейрона с сохранением полученной информации;
- влияние деполяризации, распространяемой по поверхности мембраны, на условия открытости и полуоткрытости самой мембраны по отношению к ионам калия и натрия;
- поверхностные механизмы поддержания на мембране разности потенциалов в 70-90 мВ, которой при размерах самой мембраны порядка
 [55] (
[55] (  ) соответствует напряженность электрического поля в 100000 В/см, которая более чем в 30 раз превосходит напряженность, достаточную для формирования молнии в атмосфере (напряжение "пробоя" воздуха - 3000 В/см).
) соответствует напряженность электрического поля в 100000 В/см, которая более чем в 30 раз превосходит напряженность, достаточную для формирования молнии в атмосфере (напряжение "пробоя" воздуха - 3000 В/см).
На момент создания нейроподобных вычислительных технологий (середина 50-х годов прошлого столетия) не вполне ясно было, на что больше влияет деполяризация мембраны: на механизм взвешенного суммирования или на изменение порога возбуждения. Тем не менее, впечатляющие достижения того периода в исследовании электрофизиологической активности реальных нейронов можно проиллюстрировать сводной таблицей полученных численных значений базовых структурно-параметрических характеристик нейронов (табл. 4.1).
Приведенных данных достаточно, чтобы сформулировать "системотехнический базис" реальных нейронных технологий:
- На уровне межклеточных взаимодействий в нервных сетях используются однонаправленные механизмы передачи информации по типу "цифра - аналог - цифра", что в общем случае приводит если и не к непрерывнозначной [58], то по крайней мере к многозначной [59], а не двузначной логике, правилам которой подчинены только механизмы возбуждения аксонного холмика.
- Токи в реальных нейронах и нейросетях обусловлены не столько электромагнитными взаимодействиями, сколько диффузными механизмами перемещения заряженных частиц, управление которыми осуществляется целым комплексом электрофизиологических и физико-химических факторов, изменяющих в конечном счете механические свойства полупроницаемой мембраны.
- При низких скоростях распространения "спайков" по нервному волокну темп реального времени в нервной системе и организме в целом можно поддержать комплексом методов и средств распараллеливания обработки информации и упреждающей экстраполяции как событий, происходящих во внешней среде, так и изменений собственного состояния организма.
- Условия "взвешенного" суммирования возбуждающих постсинапти-ческих потенциалов поддерживаются инактивирующим влиянием ферментов во время диффузии медиатора через синаптическую щель, изменением концентрации медиатора в зависимости от интенсивности пресинаптических возбуждений и целым рядом других достаточно случайных факторов, что в формальных моделях нейрона можно отразить только непрерывной вариаций весового и/или порогового векторов.
- Даже в двоичном базисе задание нейрону извне требуемой функции представляет собой комбинаторно неразрешимую проблему: 2^{4000}?10^{3*400}=10^{120} комбинаций значений состояний входных синапсов. Даже если рефрактерный период (период нечувствительности нейрона к входным возбуждениям - см.
рис. 4.2) составляет 0,1 мсек, то за время жизни человека (порядка 70-80 лет) на входе такого нейрона можно сформировать не более
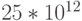 комбинаций. Это говорит о том, что поведение реального нейрона можно описать только с помощью частично определенных функций, а еще точнее - с помощью существенно недоопределенных функций.
комбинаций. Это говорит о том, что поведение реального нейрона можно описать только с помощью частично определенных функций, а еще точнее - с помощью существенно недоопределенных функций.
Таким образом, на основе приведенных данных можно заключить:
- Фундаментальные исследования физиологии синапсов [57] не столько объяснили, сколько показали всю сложность задач управления объектами с существенно недоопределенными функциями и с постоянно флуктуирующими механизмами "взвешивания" и "принятия решений", на основе которых организмы успешно синтезируют устойчивые целенаправленные формы поведения типа условных рефлексов.
- Механизмы синтеза ненадежными методами и средствами из ненадежных компонент сложных детерминированных систем являются атрибутом практически всех физиологических и нейрофизиологических исследований.